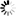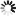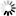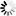Posts: 3,343
Threads: 569
Joined: Jul 2024
Reputation:
1,848
(14-08-2024, 12:21 PM)RuudVanNistelrooy Wrote: We share 80% of genes with them but we also do share many genes with bananas.
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
Quote:‘Mutations are word-processing errors in the cell’s instruction manual. Mutations systematically destroy genetic information—even as word processing errors destroy written information. While there are some rare beneficial mutations (even as there are rare beneficial misspellings),1 bad mutations outnumber them—perhaps by a million to one. So even allowing for beneficial mutations, the net effect of mutation is overwhelmingly deleterious. The more the mutations, the less the information. This is fundamental to the mutation process.’
‘Selection does help. Selection gets rid of the worst mutations. This slows mutational degeneration.
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
![[Image: pelvis.jpg]](https://dl0.creation.com/articles/p175/c17549/pelvis.jpg) Adult modern human female pelvis replica above the cast of Lucy’s reconstructed pelvis (white arrow = ilium). Note the smaller cross-sectional area of the body of Lucy’s sacrum compared to that of humans (red arrows) suggesting it was not built to bear the heavy loads associated with obligatory upright walking and/or a fully upright posture.
Adult modern human female pelvis replica above the cast of Lucy’s reconstructed pelvis (white arrow = ilium). Note the smaller cross-sectional area of the body of Lucy’s sacrum compared to that of humans (red arrows) suggesting it was not built to bear the heavy loads associated with obligatory upright walking and/or a fully upright posture.
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
Posts: 9,369
Threads: 1,235
Joined: Jul 2024
I only have trust in the Lord
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
DNA repair mechanisms ‘shout’ creation
Quote:Indeed, without the repair mechanisms, DNA falls apart “at a surprisingly fast rate”. Once an organism dies, the repair mechanisms no longer operate and DNA decays. This means that fossils that are supposedly millions of years old should not have any intact DNA; but they often do, including dinosaur fossils.
It should now be obvious to all that DNA could never have arisen by natural processes (i.e. just chemistry and physics) because even if that could happen, the DNA would be useless without the repair mechanisms to preserve it. Without the repair mechanisms, DNA would fall apart quicker than it could possibly come together (if it even could in the absence of the cellular machinery to put it together, which is itself coded on the DNA). Furthermore, no natural origin could possibly explain either the coded information storage system of the DNA or the highly ordered sequences of information on the DNA that specify how to make the protein components of life.
The repair mechanisms involve incredibly complex nano-machines3 that themselves defy the notion of naturalistic origins. The more we know about the science of life, the more it becomes clear that life came about by the action of an incredibly intelligent creator.4
![[Image: DSC05383-Editb742b4b47b71f691.jpg]](https://i.postimg.cc/cHBqm1zw/DSC05383-Editb742b4b47b71f691.jpg)
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
Quote:Given many millions of point mutations and tens of thousands of insertions, deletions, inversions, and duplications, can they explain this in an evolutionary context? They can explain some in their models, but those models are often quite simplistic (like the equations above). Random mating is a critical assumption, but it is never true, and non-random mating only slows down how fast new variants spread. There are also questions about population growth and how it affects all calculations. Given that the human population has been expanding (since the Flood or since the invention of agriculture, take your pick), ZERO genetic variants have become fixed in the human genome for the last 10,000 years in the evolutionary timeline. How does ‘no evolution for 10,000 years’ affect the evolutionary forecast?
But the fourth question above is perhaps the most fundamentally important question in evolution. Why? Because evolution needs new genes to arise and activate. Humans and chimpanzees do not just differ at the nucleotide level. Our genes are not used in the same ways and our brains have very different wiring pathways. Those changes would not just have to arise. No, they would have to arise, spread out and replace whatever original gene was in that place, and then integrate themselves into the already complex regulatory processes that exist.
The ‘fact’ that they have been trumpeting from the rooftops since the 1970s turns out to be no fact at all. The real difference is NOT 1%. No, it is 15x greater.
This is a massive problem even for the “1%” crowd.
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
The origin of life?
Being a chemist, it was almost inevitable that Royal should examine origin of life (OoL) publications. He read hundreds of chemical publications that tried to explain the OoL and concluded that “all were nothing but wishful thinking.”
For example, thousands of complex proteins are needed for cells to work. However, not one of even the simplest proteins could be created naturally. Some of the problems he noted are:
- Racemization. Nearly all the amino acids that make up proteins come in two forms, Levo (L) and Dextro (D), that are mirror images of each other like your left and right hands respectively. Only left-handed amino acids (L) make functional proteins. But they readily turn into each other, so even if you start off with pure L forms, they naturally move towards 50:50 quantities of L and D amino acids. This is called racemization. Not only do individual amino acids racemize, this happens even faster if they bond together to form proteins! Dr Truman showed that this happens so quickly under realistic conditions (temperature and pH) as to stop the formation of any functional proteins.3
- Side-chain reactions. To make functional proteins, amino acids must be joined together in the correct manner. But without the help of enzymes (which are themselves cell proteins), many will join in the wrong way.
- Reactions with other chemicals. This includes dozens of unsuitable kinds of amino acids, all the amines, all the carboxylic acids, all the alcohols, and all aldehydes. These would have dominated a hypothetical early earth. You simply cannot get the life-necessary chemicals without the ones that will interfere with the chemistry of life.
- Wrong proportions of biologically relevant amino acids. The latest research reveals that of the 20 amino acids needed for life, between 99%–99.9% produced in realistically natural processes would have been only glycine, the simplest and biologically least important amino acid.
- Obtaining long chains in water. Any experiments that join some amino acids together must remove water that is generated by the reaction, because the water hinders the joining from continuing. None having 10 or more amino acids would form in water; too short to be functional.
Dr Truman commented,
Quote:“If simply obtaining a long, linear random amino acid polymer (i.e., a ‘protein’) isn’t possible, how in the world were thousands of different proteins, each having the correct sequence of amino acids, supposed to have arisen? Natural selection can’t operate until you already have something that makes copies of itself.”
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
Another example Lents cites is the recurrent laryngeal nerve (p. 14) which he argues is much longer than required because, instead of travelling directly to the larynx, it loops around the aortic arch then travels back up to the larynx. The superior laryngeal nerve is not one nerve that has one function as implied by Lents and others, but has several functions. It divides into branches which control the internal laryngeal muscle, and innervate muscles responsible for pitch, controlling loudness, and vocal fatigue. The three main branches of the recurrent laryngeal nerve innervate several muscle bundles including the thyroarytenoid muscle, the posterior cricoarytenoid muscle and the lateral cricoarytenoid muscle. We know this because damage to the nerves that innervate these muscles affects articulation, causing speech to be ‘slurred’ or ‘garbled’. The existing recurrent laryngeal nerve system design serves to make fine adjustments to larynx control in order to improve speech quality.4
Posts: 2,696
Threads: 200
Joined: Aug 2025
Reputation:
880
Progress has not followed a straight ascending line, but a spiral with rhythms of progress and retrogression, of evolution and dissolution.
goethe
morphology>darwinism
Posts: 26,450
Threads: 3,227
Joined: Apr 2022
Reputation:
14,935
Reminder op cant tell what a cell is without googling it
(18-09-2025, 06:49 PM)MVP Wrote: i already said, rӏ is built different
Quote:![[Image: Screenshot-2026-06-20-at-23-46-52-Lookis...-Lists.png]](https://i.ibb.co/8nqCLH50/Screenshot-2026-06-20-at-23-46-52-Lookism-Edit-Buddy-and-Ignore-Lists.png)
Posts: 7,152
Threads: 711
Joined: Apr 2023
Reputation:
2,962
Quote:There’s no reason to doubt that Darwin successfully explained the small adjustments by which an organism adapts to local circumstances: changes to fur density or wing style or beak shape. Yet there are many reasons to doubt whether he can answer the hard questions and explain the big picture—not the fine-tuning of existing species but the emergence of new ones. The origin of species is exactly what Darwin cannot explain.
Stephen Meyer’s thoughtful and meticulous Darwin’s Doubt (2013) convinced me that Darwin has failed. He cannot answer the big question. Two other books are also essential: The Deniable Darwin and Other Essays (2009), by David Berlinski, and Debating Darwin’s Doubt (2015), an anthology edited by David Klinghoffer, which collects some of the arguments Meyer’s book stirred up. These three form a fateful battle group that most people would rather ignore. Bringing to bear the work of many dozen scientists over many decades, Meyer, who after a stint as a geophysicist in Dallas earned a Ph.D. in History and Philosophy of Science from Cambridge and now directs the Discovery Institute’s Center for Science and Culture, disassembles the theory of evolution piece by piece. Darwin’s Doubt is one of the most important books in a generation. Few open-minded people will finish it with their faith in Darwin intact.
Meyer doesn’t only demolish Darwin; he defends a replacement theory, intelligent design (I.D.). Although I can’t accept intelligent design as Meyer presents it, he does show that it is a plain case of the emperor’s new clothes: it says aloud what anyone who ponders biology must think, at some point, while sifting possible answers to hard questions. Intelligent design as Meyer explains it never uses religious arguments, draws religious conclusions, or refers to religion in any way. It does underline an obvious but important truth: Darwin’s mission was exactly to explain the flagrant appearance of design in nature.
The religion is all on the other side. Meyer and other proponents of I.D. are the dispassionate intellectuals making orderly scientific arguments. Some I.D.-haters have shown themselves willing to use any argument—fair or not, true or not, ad hominem or not—to keep this dangerous idea locked in a box forever. They remind us of the extent to which Darwinism is no longer just a scientific theory but the basis of a worldview, and an emergency replacement religion for the many troubled souls who need one.
As for Biblical religion, it forces its way into the discussion although Meyer didn’t invite it, and neither did Darwin. Some have always been bothered by the harm Darwin is said to have done religion. His theory has been thought by some naïfs (fundamentalists as well as intellectuals) to have shown or alleged that the Bible is wrong, and Judeo-Christian religion bunk. But this view assumes a childishly primitive reading of Scripture. Anyone can see that there are two different creation stories in Genesis, one based on seven days, the other on the Garden of Eden. When the Bible gives us two different versions of one story, it stands to reason that the facts on which they disagree are without basic religious significance. The facts on which they agree are the ones that matter: God created the universe, and put man there for a reason. Darwin has nothing to say on these or any other key religious issues.
Fundamentalists and intellectuals might go on arguing these things forever. But normal people will want to come to grips with Meyer and the downfall of a beautiful idea. I will mention several of his arguments, one of them in (just a bit of) detail. This is one of the most important intellectual issues of modern times, and every thinking person has the right and duty to judge for himself.
Looking for Evidence
Darwin himself had reservations about his theory, shared by some of the most important biologists of his time. And the problems that worried him have only grown more substantial over the decades. In the famous “Cambrian explosion” of around half a billion years ago, a striking variety of new organisms—including the first-ever animals—pop up suddenly in the fossil record over a mere 70-odd million years. This great outburst followed many hundreds of millions of years of slow growth and scanty fossils, mainly of single-celled organisms, dating back to the origins of life roughly three and half billion years ago.
Darwin’s theory predicts that new life forms evolve gradually from old ones in a constantly branching, spreading tree of life. Those brave new Cambrian creatures must therefore have had Precambrian predecessors, similar but not quite as fancy and sophisticated. They could not have all blown out suddenly, like a bunch of geysers. Each must have had a closely related predecessor, which must have had its own predecessors: Darwinian evolution is gradual, step-by-step. All those predecessors must have come together, further back, into a series of branches leading down to the (long ago) trunk.
But those predecessors of the Cambrian creatures are missing. Darwin himself was disturbed by their absence from the fossil record. He believed they would turn up eventually. Some of his contemporaries (such as the eminent Harvard biologist Louis Agassiz) held that the fossil record was clear enough already, and showed that Darwin’s theory was wrong. Perhaps only a few sites had been searched for fossils, but they had been searched straight down. The Cambrian explosion had been unearthed, and beneath those Cambrian creatures their Precambrian predecessors should have been waiting—and weren’t. In fact, the fossil record as a whole lacked the upward-branching structure Darwin predicted.
The trunk was supposed to branch into many different species, each species giving rise to many genera, and towards the top of the tree you would find so much diversity that you could distinguish separate phyla—the large divisions (sponges, mosses, mollusks, chordates, and so on) that comprise the kingdoms of animals, plants, and several others—take your pick. But, as Berlinski points out, the fossil record shows the opposite: “representatives of separate phyla appearing first followed by lower-level diversification on those basic themes.” In general, “most species enter the evolutionary order fully formed and then depart unchanged.” The incremental development of new species is largely not there. Those missing pre-Cambrian organisms have still not turned up. (Although fossils are subject to interpretation, and some biologists place pre-Cambrian life-forms closer than others to the new-fangled Cambrian creatures.)
Some researchers have guessed that those missing Precambrian precursors were too small or too soft-bodied to have made good fossils. Meyer notes that fossil traces of ancient bacteria and single-celled algae have been discovered: smallness per se doesn’t mean that an organism can’t leave fossil traces—although the existence of fossils depends on the surroundings in which the organism lived, and the history of the relevant rock during the ages since it died. The story is similar for soft-bodied organisms. Hard-bodied forms are more likely to be fossilized than soft-bodied ones, but many fossils of soft-bodied organisms and body parts do exist. Precambrian fossil deposits have been discovered in which tiny, soft-bodied embryo sponges are preserved—but no predecessors to the celebrity organisms of the Cambrian explosion.
This sort of negative evidence can’t ever be conclusive. But the ever-expanding fossil archives don’t look good for Darwin, who made clear and concrete predictions that have (so far) been falsified—according to many reputable paleontologists, anyway. When does the clock run out on those predictions? Never. But any thoughtful person must ask himself whether scientists today are looking for evidence that bears on Darwin, or looking to explain away evidence that contradicts him. There are some of each. Scientists are only human, and their thinking (like everyone else’s) is colored by emotion.
The Advent of Molecular Biology
Darwin’s main problem, however, is molecular biology. There was no such thing in his own time. We now see from inside what he could only see from outside, as if he had developed a theory of mobile phone evolution without knowing that there were computers and software inside or what the digital revolution was all about. Under the circumstances, he did brilliantly.
Biology in his time was for naturalists, not laboratory scientists. Doctor Dolittle was a naturalist. (He is the hero of the wonderful children’s books by Hugh Lofting, now unfortunately nearing extinction.) The doctor loved animals and understood them, and had a sharp eye for all of nature not too different from Wordsworth’s or Goethe’s. But the character of the field has changed, and it’s not surprising that old theories don’t necessarily still work.
Darwin’s theory is simple to grasp; its simplicity is the heart of its brilliance and power. We all know that variation occurs naturally among individuals of the same type—white or black sheep, dove-gray versus off-white or pale beige pigeons, boring and sullen undergraduates versus charming, lissome ones. We all know that many variations have no effect on a creature’s prospects, but some do. A sheep born with extra-warm wool will presumably do better at surviving a rough Scottish winter than his normal-wooled friends. Such a sheep would be more likely than normal sheep to live long enough to mate, and pass on its superior trait to the next generation. Over millions of years, small good-for-survival variations accumulate, and eventually (says Darwin) you have a brand new species. The same mechanism naturally favors genes that are right for the local environment—warm wool in Scotland, light and comfortable wool for the tropics, other varieties for mountains and deserts. Thus one species (your standard sheep) might eventually become four specialized ones. And thus new species should develop from old in the upward-branching tree pattern Darwin described.
The advent of molecular biology made it possible to transform Darwinism into Neo-Darwinism. The new version explains (it doesn’t merely cite) natural variation, as the consequence of random change or mutation to the genetic information within cells that deal with reproduction. Those cells can pass genetic change onward to the next generation, thus changing—potentially—the future of the species and not just one individual’s career.
The engine that powers Neo-Darwinian evolution is pure chance and lots of time. By filling in the details of cellular life, molecular biology makes it possible to estimate the power of that simple mechanism. But what does generating new forms of life entail? Many biologists agree that generating a new shape of protein is the essence of it. Only if Neo-Darwinian evolution is creative enough to do that is it capable of creating new life-forms and pushing evolution forward.
Proteins are the special ops forces (or maybe the Marines) of living cells, except that they are common instead of rare; they do all the heavy lifting, all the tricky and critical assignments, in a dazzling range of roles. Proteins called enzymes catalyze all sorts of reactions and drive cellular metabolism. Other proteins (such as collagen) give cells shape and structure, like tent poles but in far more shapes. Nerve function, muscle function, and photosynthesis are all driven by proteins. And in doing these jobs and many others, the actual, 3-D shape of the protein molecule is important.
So, is the simple neo-Darwinian mechanism up to this task? Are random mutation plus natural selection sufficient to create new protein shapes?
Mutations
How to make proteins is our first question. Proteins are chains: linear sequences of atom-groups, each bonded to the next. A protein molecule is based on a chain of amino acids; 150 elements is a “modest-sized” chain; the average is 250. Each link is chosen, ordinarily, from one of 20 amino acids. A chain of amino acids is a polypeptide—“peptide” being the type of chemical bond that joins one amino acid to the next. But this chain is only the starting point: chemical forces among the links make parts of the chain twist themselves into helices; others straighten out, and then, sometimes, jackknife repeatedly, like a carpenter’s rule, into flat sheets. Then the whole assemblage folds itself up like a complex sheet of origami paper. And the actual 3-D shape of the resulting molecule is (as I have said) important.
Imagine a 150-element protein as a chain of 150 beads, each bead chosen from 20 varieties. But: only certain chains will work. Only certain bead combinations will form themselves into stable, useful, well-shaped proteins.
So how hard is it to build a useful, well-shaped protein? Can you throw a bunch of amino acids together and assume that you will get something good? Or must you choose each element of the chain with painstaking care? It happens to be very hard to choose the right beads.
Inventing a new protein means inventing a new gene. (Enter, finally, genes, DNA etc., with suitable fanfare.) Genes spell out the links of a protein chain, amino acid by amino acid. Each gene is a segment of DNA, the world’s most admired macromolecule. DNA, of course, is the famous double helix or spiral staircase, where each step is a pair of nucleotides. As you read the nucleotides along one edge of the staircase (sitting on one step and bumping your way downwards to the next and the next), each group of three nucleotides along the way specifies an amino acid. Each three-nucleotide group is a codon, and the correspondence between codons and amino acids is the genetic code. (The four nucleotides in DNA are abbreviated T, A, C and G, and you can look up the code in a high school textbook: TTA and TTC stand for phenylalanine, TCT for serine, and so on.)
Your task is to invent a new gene by mutation—by the accidental change of one codon to a different codon. You have two possible starting points for this attempt. You could mutate an existing gene, or mutate gibberish. You have a choice because DNA actually consists of valid genes separated by long sequences of nonsense. Most biologists think that the nonsense sequences are the main source of new genes. If you tinker with a valid gene, you will almost certainly make it worse—to the point where its protein misfires and endangers (or kills) its organism—long before you start making it better. The gibberish sequences, on the other hand, sit on the sidelines without making proteins, and you can mutate them, so far as we know, without endangering anything. The mutated sequence can then be passed on to the next generation, where it can be mutated again. Thus mutations can accumulate on the sidelines without affecting the organism. But if you mutate your way to an actual, valid new gene, your new gene can create a new protein and thereby, potentially, play a role in evolution.
Mutations themselves enter the picture when DNA splits in half down the center of the staircase, thereby allowing the enclosing cell to split in half, and the encompassing organism to grow. Each half-staircase summons a matching set of nucleotides from the surrounding chemical soup; two complete new DNA molecules emerge. A mistake in this elegant replication process—the wrong nucleotide answering the call, a nucleotide typo—yields a mutation, either to a valid blueprint or a stretch of gibberish.
Building a Better Protein
Now at last we are ready to take Darwin out for a test drive. Starting with 150 links of gibberish, what are the chances that we can mutate our way to a useful new shape of protein? We can ask basically the same question in a more manageable way: what are the chances that a random 150-link sequence will create such a protein? Nonsense sequences are essentially random. Mutations are random. Make random changes to a random sequence and you get another random sequence. So, close your eyes, make 150 random choices from your 20 bead boxes and string up your beads in the order in which you chose them. What are the odds that you will come up with a useful new protein?
It’s easy to see that the total number of possible sequences is immense. It’s easy to believe (although non-chemists must take their colleagues’ word for it) that the subset of useful sequences—sequences that create real, usable proteins—is, in comparison, tiny. But we must know how immense and how tiny.
The total count of possible 150-link chains, where each link is chosen separately from 20 amino acids, is 20150. In other words, many. 20150 roughly equals 10195, and there are only 1080 atoms in the universe.
What proportion of these many polypeptides are useful proteins? Douglas Axe did a series of experiments to estimate how many 150-long chains are capable of stable folds—of reaching the final step in the protein-creation process (the folding) and of holding their shapes long enough to be useful. (Axe is a distinguished biologist with five-star breeding: he was a graduate student at Caltech, then joined the Centre for Protein Engineering at Cambridge. The biologists whose work Meyer discusses are mainly first-rate Establishment scientists.) He estimated that, of all 150-link amino acid sequences, 1 in 1074 will be capable of folding into a stable protein. To say that your chances are 1 in 1074 is no different, in practice, from saying that they are zero. It’s not surprising that your chances of hitting a stable protein that performs some useful function, and might therefore play a part in evolution, are even smaller. Axe puts them at 1 in 1077.
In other words: immense is so big, and tiny is so small, that neo-Darwinian evolution is—so far—a dead loss. Try to mutate your way from 150 links of gibberish to a working, useful protein and you are guaranteed to fail. Try it with ten mutations, a thousand, a million—you fail. The odds bury you. It can’t be done.
A Bad Bet
But neo-Darwinianism understands that mutations are rare, and successful ones even scarcer. To balance that out, there are many organisms and a staggering immensity of time. Your chances of winning might be infinitesimal. But if you play the game often enough, you win in the end, right? After all, it works for Powerball!
Do the numbers balance out? Is Neo-Darwinian evolution plausible after all? Axe reasoned as follows. Consider the whole history of living things—the entire group of every living organism ever. It is dominated numerically by bacteria. All other organisms, from tangerine trees to coral polyps, are only a footnote. Suppose, then, that every bacterium that has ever lived contributes one mutation before its demise to the history of life. This is a generous assumption; most bacteria pass on their genetic information unchanged, unmutated. Mutations are the exception. In any case, there have evidently been, in the whole history of life, around 1040 bacteria—yielding around 1040 mutations under Axe’s assumptions. That is a very large number of chances at any game. But given that the odds each time are 1 to 1077 against, it is not large enough. The odds against blind Darwinian chance having turned up even one mutation with the potential to push evolution forward are 1040x(1/1077)—1040 tries, where your odds of success each time are 1 in 1077—which equals 1 in 1037. In practical terms, those odds are still zero. Zero odds of producing a single promising mutation in the whole history of life. Darwin loses.
His idea is still perfectly reasonable in the abstract. But concretely, he is overwhelmed by numbers he couldn’t possibly have foreseen: the ridiculously large number of amino-acid chains relative to number of useful proteins. Those numbers transcend the details of any particular set of estimates. The obvious fact is that genes, in storing blueprints for the proteins that form the basis of cellular life, encode an awe-inspiring amount of information. You don’t turn up a useful protein merely by doodling on the back of an envelope, any more than you write a Mozart aria by assembling three sheets of staff paper and scattering notes around. Profound biochemical knowledge is somehow, in some sense, captured in every description of a working protein. Where on earth did it all come from?
Neo-Darwinianism says that nature simply rolls the dice, and if something useful emerges, great. Otherwise, try again. But useful sequences are so gigantically rare that this answer simply won’t work. Studies of the sort Meyer discusses show that Neo-Darwinism is the quintessence of a bad bet.
The Great Darwinian Paradox
There are many other problems besides proteins. One of the most basic, and the last I’ll mention here, calls into question the whole idea of gene mutations driving macro-evolution—the emergence of new forms of organism, versus mere variation on existing forms.
To help create a brand new form of organism, a mutation must affect a gene that does its job early and controls the expression of other genes that come into play later on as the organism grows. But mutations to these early-acting “strategic” genes, which create the big body-plan changes required by macro-evolution, seem to be invariably fatal. They kill off the organism long before it can reproduce. This is common sense. Severely deformed creatures don’t ever seem fated to lead the way to glorious new forms of life. Instead, they die young.
Evidently there are a total of no examples in the literature of mutations that affect early development and the body plan as a whole and are not fatal. The German geneticists Christiane Nüsslein-Volhard and Eric Wieschaus won the Nobel Prize in 1995 for the “Heidelberg screen,” an exhaustive investigation of every observable or inducible mutation of Drosophila melanogaster (the same patient, long-suffering fruit fly I meddled with relentlessly in an undergraduate genetics lab in the 1970s). “[W]e think we’ve hit all the genes required to specify the body plan of Drosophila,” said Wieschaus in answering a question after a talk. Not one, he continued, is “promising as raw materials for macroevolution”—because mutations in them all killed off the fly long before it could mate. If an exhaustive search rules out every last plausible gene as a candidate for large-scale Drosophila evolution, where does that leave Darwin? Wieschaus continues: “What are—or what would be—the right mutations for major evolutionary change? And we don’t know the answer to that.”
There is a general principle here, similar to the earlier principle that the number of useless polypeptides crushes the number of useful ones. The Georgia Tech geneticist John F. McDonald calls this one a “great Darwinian paradox.” Meyer explains: “genes that are obviously variable within natural populations seem to affect only minor aspects of form and function—while those genes that govern major changes, the very stuff of macroevolution, apparently do not vary or vary only to the detriment of the organism.” The philosopher of biology Paul Nelson summarizes the body-plan problem:
Research on animal development and macroevolution over the last thirty years—research done from within the neo-Darwinian framework—has shown that the neo-Darwinian explanation for the origin of new body plans is overwhelmingly likely to be false—and for reasons that Darwin himself would have understood.
Darwin would easily have understood that minor mutations are common but can’t create significant evolutionary change; major mutations are rare and fatal.
Posts: 5,393
Threads: 210
Joined: Apr 2022
Reputation:
2,560
(07-09-2025, 09:41 PM)Altruist Wrote: Reminder op cant tell what a cell is without googling it Enlighten me about the cell but without a microscope.
Posts: 7,152
Threads: 711
Joined: Apr 2023
Reputation:
2,962
(07-09-2025, 09:34 PM)machinelves Wrote: Progress has not followed a straight ascending line, but a spiral with rhythms of progress and retrogression, of evolution and dissolution.
goethe
morphology>darwinism
![[Image: 213124124.png]](https://i.ibb.co/84b2D8fv/213124124.png)
Posts: 7,152
Threads: 711
Joined: Apr 2023
Reputation:
2,962
|


![[Image: 214214124.png]](https://i.ibb.co/zjzTjrD/214214124.png)
![[Image: paracelsus_pansophic_conception_universe_1618.jpg]](https://www.sciencehistory.org/wp-content/uploads/2023/04/paracelsus_pansophic_conception_universe_1618.jpg)